 I suspect I’m one of many for which the predictability of species distributions was a gateway drug to ecology. My journey began with an extraordinary biogeographic transition point, delimited by the sheer slopes of the Plenty Gorge on Melbourne’s northern outskirts. East of the river – where I grew up – was a world of Yellow Gum, Red Box, Wattle and Cassinia. Undulating mile upon undulating mile of it, built on sedimentary soils laid down during the Ordovician. The plains west of the Plenty River were a very different landscape. Volcanic epochs transformed this country, laying deep basalt clays over the underlying sedimentary rock. Kangaroo and Wallaby Grass, Spear Grass and Poa grew here in prolific stretches prior to European settlement, and gave rise to a markedly different faunal assemblage to that east of the Gorge.
I suspect I’m one of many for which the predictability of species distributions was a gateway drug to ecology. My journey began with an extraordinary biogeographic transition point, delimited by the sheer slopes of the Plenty Gorge on Melbourne’s northern outskirts. East of the river – where I grew up – was a world of Yellow Gum, Red Box, Wattle and Cassinia. Undulating mile upon undulating mile of it, built on sedimentary soils laid down during the Ordovician. The plains west of the Plenty River were a very different landscape. Volcanic epochs transformed this country, laying deep basalt clays over the underlying sedimentary rock. Kangaroo and Wallaby Grass, Spear Grass and Poa grew here in prolific stretches prior to European settlement, and gave rise to a markedly different faunal assemblage to that east of the Gorge.
As a young fellow, this biogeographic divide enabled an idea to crystallise in my mind. If I could grasp the geological, climatic or floristic affiliations of species, I could always find them. Little Whip Snake? West of the river, on cracking clay soils under exfoliating basalt. Weasel Skink? Rotting leaf litter or timber in damp gullies east of the river. Jacky Dragon? Cassinia and Acacia dominated heathlands on the steeper banks of the Gorge, where protruding mudstone reefs dominate. These were rules, as good as the laws of thermodynamics – learn them, and you have each species pegged.
But what if you don’t? What if these rules can change? What if much of what you have learnt about a species can go out the window in a metaphoric blink of the eye? Welcome to the concept of the ‘dynamic niche’, where biotic shifts can drastically reshape the distribution, habitat affiliations, demography and even evolutionary trajectory of species.
Over recent decades, amphibian ecologists have been confronted with rapid niche shifts of this kind. The driver has been the emergence and spread of the pandemic lineage of chytrid fungus. The headline grabbing impacts of this pathogen have been wholesale extinction events and drastic population declines. But another, more subtle phenomenon has also emerged – changes in the ecology of many frogs, some of which now appear quite irreversible. Our most recent paper, led by Ben Scheele, is an attempt to describe these shifts, to synthesise the patterns therein, and to guide conservation responses for species that appear permanently changed by their interactions with this pathogen.
Two frog species from Australia’s south-east are useful exemplars. The first is my beloved Growling Grass Frog, a species that was both abundant and widespread prior to the arrival of chytrid to Australia’s east coast in the late 1970s. Through the 1980s and 1990s, the species experienced drastic population declines, collapsing out of the areas that were environmentally most suitable for the pathogen – colder and/or wetter parts of the range, such as higher-elevations, frost-ridden plains and wetter foothills (see figure below). Habitat affiliations of the frog also appear to have narrowed, to wetlands that are relatively warm and/or slightly salt-affected (conditions in which the pathogenicity and virulence of chytrid is reduced). No longer can they persist in the cooler, fresher sites in which they once thrived, nor those that dry out with any frequency. The Growling Grass Frog we know today is a different beast to the frog we knew prior to 1978.
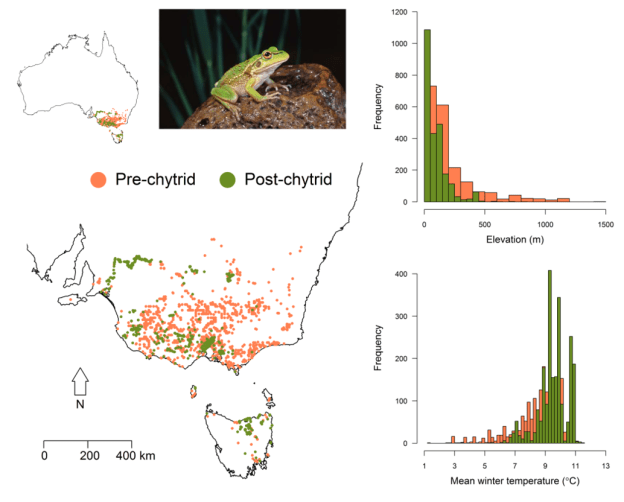
Changes in the distribution and realised niche of the Growling Grass Frog due to chytrid. The map shows records of the species up to 1980 (orange) and subsequent to 2000 (green), broadly representing the pre- and post-chytrid distribution. The plots show the change in the elevational and thermal niche of the species, resulting from its distributional contraction. Figure 2 from Scheele et al. (2019), Biological Conservation 236, 52-59. © Elsevier.
Likewise, the Alpine Tree Frog has been changed dramatically by the arrival of chytrid. A once fairly long-lived species (up to 7 years), today it rarely lives beyond 2 years of age. Early reproduction is now imperative, and there is evidence the heavy selection for early reproduction is altering the life history traits of this species. Habitat affiliations are also shifting, because consistent recruitment is now utterly vital for persistence at the population level (with adult survival so low, even a single year of reproductive failure can spell doom for isolated populations). As a result, the species is dropping out of the ephemeral wetlands systems that were once prime breeding habitat, and it is increasingly affiliated with permanent or near-permanent wetlands.
As well as highlighting the ecological impacts of chytrid, these two species are useful for demonstrating the shift that conservation managers must make in their approach to species with endemic chytrid infections. Take re-introduction to former haunts, which is often proposed as an approach to conserve these frogs and many others. Such initiatives will fail miserably if chytrid has the upper hand at the target locations. Indeed, we’ve seen this for another Australian hylid for which re-introduction has been attempted numerous times – the Green and Golden Bell Frog. Most re-introductions have failed, often as the frogs were put into habitat with characteristics that they used to be affiliated with. Ultimately, these re-introductions failed because we, as amphibian ecologists, failed to recognise the new world order for these frogs.
For more on the topic, you can find the paper here (or email me for a copy if you can’t get through the paywall). It is a thought-provoking, challenging and perhaps even controversial concept, but one we must consider for effective amphibian conservation in the post-chytrid era. Nevertheless, the parallels with invasive species impacts and wildlife diseases more generally are clear, and so we hope the paper will prove of interest beyond the realms of amphibian conservation. Niche shifts are happening across the globe, and we must be alert to them if our conservation initiatives are to remain effective.
