
Blackops. There are few better feelings. Midnight is long past and the moon is at is zenith. No need for headlamp as you cut across the reserve. You’ve been smart, parking a good kilometre away, and your approach has been flawless. Well, almost flawless. The distant uproar of a Doberman and the scattering Eastern Greys have given you moments to pause and bunker down in the weeds. Those clued in to the stirrings of the night notice such things. But silence returns and it’s time to make for the escarpment. Find the ‘roo track, which cuts down through the basalt. The entrance is hidden among the Gorse, remember? As you descend into the gorge, and to seclusion, you can finally turn on your headlamp. Your quarry is close. There. In the reeds. Eyeshine.
I’ve spent countless nights like this over the years, sculking around the creeks and wetlands of northern Melbourne in search of frogs. It’s a special experience to be the only soul around, quietly documenting an extraordinary ecological phenomenon. A frog’s battle for survival in a landscape it has occupied for thousands – and possibly millions – of years. Which species? The Growling Grass Frog, of course, to which I’ve been slightly addicted for going on 25 years. These stunning green and golden frogs are under siege around Melbourne.
In the late 1970s, a fungal pathogen invaded our shores. It spread rapidly across south-eastern Australia, with devastating impacts on the region’s frog fauna. This fungus – known to the legions as ‘chytrid fungus’ – had a unique capacity to devour frog skin, eating away the keratin that holds it together and rapidly causing death for highly susceptible frogs. Our stunning green amphibians of the genus Ranoidea died like flies.
But not everywhere. The outskirts of Melbourne, strangely, were a refuge. Like certain other areas within the species’ range, the climatic and geological characteristics of northern and western Melbourne were not to the fungus’ liking. Too warm, too dry and with volcanic soils that ensured many wetlands are slightly brackish – with modest salinity and high pH. Conditions in which the frogs had the upper hand and managed to cling to existence wherever wetlands with the right characteristics were found.
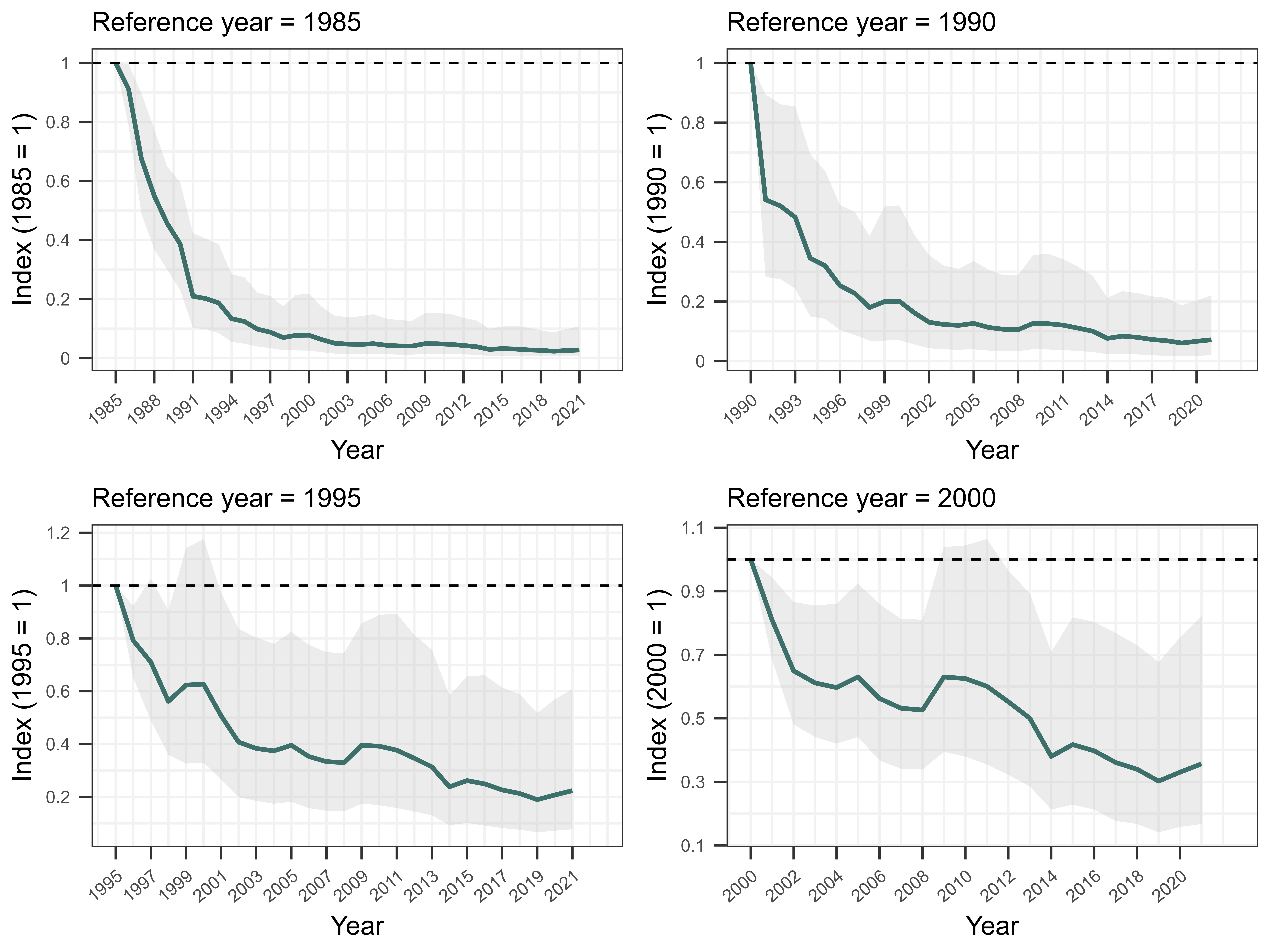
In my first season working on Growling Grass Frogs – the summer of 2001/2002 – such wetland networks were plentiful across northern Melbourne. And the frogs along with them. But two things have played out since then, which we’ve documented over two decades of monitoring. The first is that when the surveys started, chytrid hadn’t quite snuffed out all the populations it was eventually going to. To explain, we must briefly dig into the world of extinction lags. An extinction lag is a delay in the recalibration of species populations when they face a new threat. All biological systems have some inertia, and extinction lags are the time it takes for a population to succumb to a threat it cannot, in the long-term, survive. For frogs battling chytrid fungus, extinction lags arise because birth and death rates wobble around from year to year, and even when recruitment is not sufficient to replace chytrid-driven mortality in the long-term, some populations can outrun the fungus for a while, if conditions are good. This proved true for Growlers around northern Melbourne, with numerous populations that we found in 2001/2002 blipping out shortly thereafter, when recruitment failed and chytrid finally dealt the fatal blow.
The second thing that has occurred across northern Melbourne since that first year of surveys is massive urban expansion. The region is a designated urban growth corridor. This has significant implications for Growlers. Wetlands get drained and destroyed, streams become polluted and devoid of aquatic vegetation, and houses, factories, roads and rail-lines cut off dispersal corridors between key habitat patches.
Two major blows. But, what 20 odd years of monitoring data reveal, is that these two threats don’t operate independently to drive population declines. In our new paper, we show that chytrid fungus and urbanisation in fact appear to operate synergistically to extirpate Growler populations. By coupling statistical models of the occupancy dynamics of Growlers with simulations that isolate the impacts of each threat, we show that the combined impacts of chytrid fungus and urbanisation are bigger than the sum of their parts.
Why? Well, as our previous research has shown, chytrid-induced mortality leaves Growler populations on a knife edge. They rely on high and consistent recruitment to outrun deaths from infections. One or two bad years with few recruits and the populations are doomed. Urbanisation makes this more likely, by changing the hydrology of streams and wetlands, causing pollution through stormwater inputs, and degrading the aquatic vegetation that is vital for egg and tadpole survival. Urbanisation also cuts populations off from each other. In turn, populations that suffer recruitment failure cannot be rescued by recruits from a neighbouring population, and recolonisation following a local extinction event is also much less likely. Taken together, chytrid and urbanisation unravel the natural resilience Growler populations have to environmental vagaries they can otherwise overcome. And so, under these interacting threats, Growler populations across northern Melbourne have declined in number by 38% since 2001/2002.
This coming season – 2026/2027 – marks 25 years since we started monitoring Growlers across northern Melbourne. Blackops are firmly back on the cards. But what will we find? Urbanisation has marched on relentlessly since our last full round of surveys in 2021/2022. I fear the decline will only have hastened, and so many former haunts of the species will now be silent. But I can also dare to dream that some recovery may be evident. In 2021/2022, cold and wet La Niña conditions were proving the perfect conditions for chytrid, and Growlers were absent from sites that they just really shouldn’t have been. We know already that some Growler populations have made it through, and I can only hope that the intervening years have seen an amphibious insurgency against the dual enemies of chytrid and urban expansion.










 Cold, wet, exhausted. It’s 2AM and you’re 1000 m up a rainforest ravine in the Wet Tropics. You’re finally horizontal, but your hammock – saturated and starting to smell of mildew – offers little prospect of sleep. Outside, in the pitch black, the stream you’ve been relentlessly climbing cascades over boulders that it has been wearing down for millennia. But over the white noise of water on granite, you can hear a frog calling “wreek wreek wreek”. It’s the Common Mistfrog, and you’ve just detected the species high atop the jungle-clad Mt Lewis. In the process, you’ve confirmed that this species is winning the battle against
Cold, wet, exhausted. It’s 2AM and you’re 1000 m up a rainforest ravine in the Wet Tropics. You’re finally horizontal, but your hammock – saturated and starting to smell of mildew – offers little prospect of sleep. Outside, in the pitch black, the stream you’ve been relentlessly climbing cascades over boulders that it has been wearing down for millennia. But over the white noise of water on granite, you can hear a frog calling “wreek wreek wreek”. It’s the Common Mistfrog, and you’ve just detected the species high atop the jungle-clad Mt Lewis. In the process, you’ve confirmed that this species is winning the battle against 

 Rainforest glamping? Not at 1000 m up a ravine, with blood oozing from myriad leech bites, battered shins and little more than gluggy porridge awaiting for breakfast.
Rainforest glamping? Not at 1000 m up a ravine, with blood oozing from myriad leech bites, battered shins and little more than gluggy porridge awaiting for breakfast. I suspect I’m one of many for which the predictability of species distributions was a gateway drug to ecology. My journey began with an extraordinary biogeographic transition point, delimited by the sheer slopes of the
I suspect I’m one of many for which the predictability of species distributions was a gateway drug to ecology. My journey began with an extraordinary biogeographic transition point, delimited by the sheer slopes of the 
 Road trips of my youth rarely entailed music. My siblings and I cruised the highways of south-eastern Australia mostly in silence. Melbourne to Bright – silence. Melbourne to Barmah – silence. Melbourne to Canberra – silence. Melbourne to Mildura – silence.
Road trips of my youth rarely entailed music. My siblings and I cruised the highways of south-eastern Australia mostly in silence. Melbourne to Bright – silence. Melbourne to Barmah – silence. Melbourne to Canberra – silence. Melbourne to Mildura – silence. Last week myself and colleagues from Charles Sturt University, Deakin University and the University of Queensland wrote a piece for
Last week myself and colleagues from Charles Sturt University, Deakin University and the University of Queensland wrote a piece for  How does one control a rapacious pathogen? If it were an infectious agent of humans, we would have much in our armoury. We could
How does one control a rapacious pathogen? If it were an infectious agent of humans, we would have much in our armoury. We could 

